海马体联想记忆的理论及模型实验,对整个海马-新皮质区进行建模

海马体联想记忆的理论及模型实验,对整个海马-新皮质区进行建模

CreateAMind
发布于 2023-09-13 20:37:02
发布于 2023-09-13 20:37:02
论文选读:

先上理论和公式图表


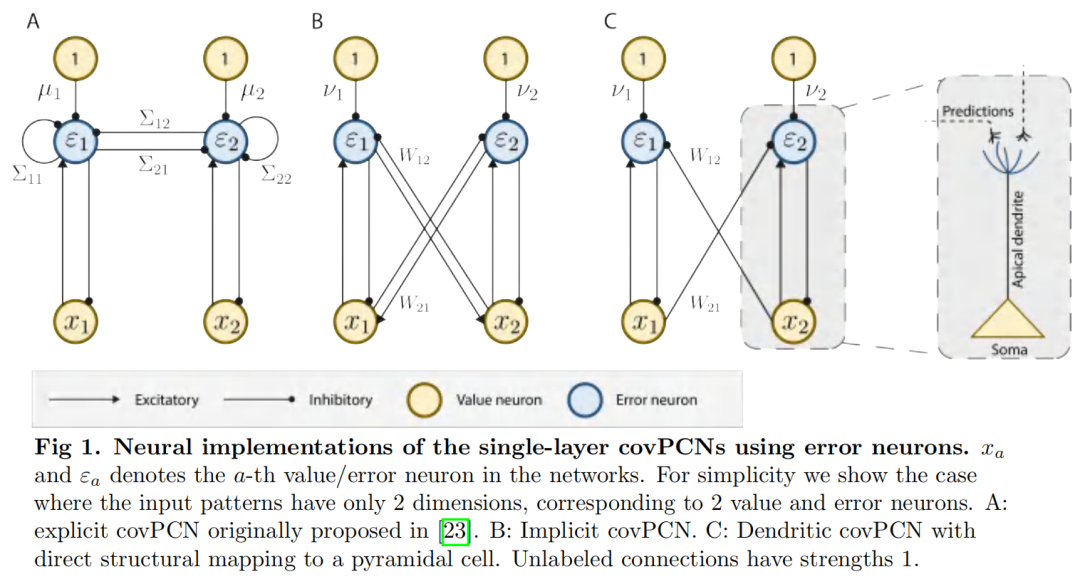

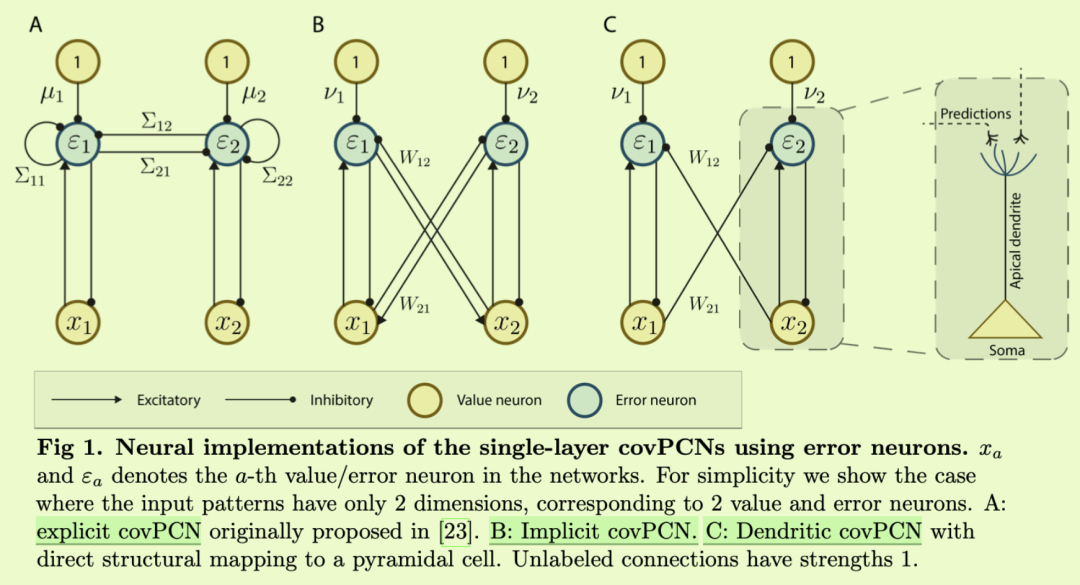
摘要
海马在联想记忆( associative memory AM)任务中采用的计算原则一直是计算和理论神经科学中最主要的研究课题之一。海马网络的经典模型假设AM是通过一种形式的协方差学习来执行的,其中记忆项目之间的关联由学习的协方差矩阵中的条目来表示,该学习的协方差矩阵编码在海马子场CA3中的循环连接中。另一方面,最近有人提出,海马中的AM是通过预测编码实现的。遵循这一理论的分级预测编码模型执行AM,但未能捕获编码经典模型中协方差的递归海马结构。这种二分法对发展记忆如何在海马体中形成和回忆的统一理论造成了潜在的困难。早期的预测编码模型明确地学习输入的协方差信息,似乎是这种二分法的解决方案。在这里,我们表明,尽管这些模型可以执行AM,但它们是以一种不可信和数值不稳定的方式执行的。相反,我们提出了这些早期协方差学习预测编码网络的替代方案,这些网络隐式地和似是而非地学习协方差信息,并可以使用树枝状结构来编码预测误差。我们通过分析表明,我们提出的模型完全等价于早期的预测编码模型学习协方差,并且在实际执行AM任务时不会遇到数值问题。我们进一步表明,我们的模型可以与分层预测编码网络相结合,以模拟海马-新皮质的相互作用。我们的模型提供了一种生物学上可行的方法来模拟海马网络,指出了海马在记忆形成和回忆过程中使用的潜在计算机制,该机制基于递归网络结构统一了预测编码和协方差学习。
作者摘要
海马和邻近的皮质区域长期以来被认为是形成联想记忆的关键。早期的理论工作假设海马体在它的循环连接中储存了嵌入在感觉输入中的统计规则。另一方面,最近有人提出,海马通过预测正在进行的感觉输入来检索记忆。
因此,计算模型被提出来解释海马网络使用预测编码的预测性质,预测编码是大脑皮层信息处理的一般理论。然而,海马体的这些分层预测编码模型没有描述它如何存储在经典海马体模型中对联想记忆起关键作用的统计规律,这阻碍了对海马体所采用的潜在计算原理的统一理解。为了解决这种二分法,这里我们提出了一个预测编码模型家族,它也学习联想记忆所需的统计信息。我们的模型可以以一种生物学上看似合理的方式稳定地执行联想记忆任务,即使是对大型结构化数据,如自然场景。我们的工作提供了一种可能的机制,即反复出现的海马网络如何同时利用各种计算原理来执行联想记忆。
介绍
大脑中的记忆系统通常存储关于对象或概念之间的关系或关联的信息。这种特殊类型的记忆,被称为联想记忆(AM),在我们的日常生活中无处不在。例如,我们会记住某个特定品牌香水的味道,一种咖啡的味道,或者我们喜欢的不同歌手的声音。在记忆形成后,当相关的线索呈现给我们的感官时,AM将利用所提供的线索和缺失成分之间的学习关联来支持其检索。
长期以来,人们一直认为位于大脑内侧颞叶的海马体和邻近的皮质区对AM至关重要[1-3]。同时,理论和计算工作已经发展到在AM任务中努力模拟海马体[4,5]。沿着这个方向的早期工作,如Hopfield网络[6],稀疏分布存储器[7,8]和相关矩阵存储器[9],集中于简单和随机模式的存储和检索。这些模型的简单性也允许对它们的容量和检索保真度进行广泛的理论研究[10,11]。然而,这些网络模型通常无法记忆和检索自然图像等结构化数据,这些模型的现代变体已被提出来解决这一问题[12–14]。这些模型共有的一个共同特性是,它们假设海马网络的循环连接通过对协方差矩阵进行编码来支持输入模式的记忆,协方差矩阵代表不同神经元之间的活动如何共同变化。
协方差矩阵的元素代表由记忆项目激活的单个神经元之间的关联,从而形成关联记忆,然后可以用于在检索过程中回忆起记忆中缺失的成分。
另一项研究遵循了海马能够预测正在进行的感觉输入的实验发现[15,16],并且提出了描述海马如何通过预测处理存储和恢复记忆的理论预测编码(PC)框架[17]。Rao [18]在开创性的工作中已经证明了预测编码网络(PCN)完成先前学习的模式的能力。最近,Salvatori等人对这些网络充当AM的能力进行了计算分析[19]。在这项工作中,作者训练了最初由Rao和Ballard [20]开发的纯层次PCN,以模拟海马体-新皮质的相互作用,并表明这些网络可以将训练数据点存储为记忆,并在给定部分或有噪声的线索的情况下检索这些记忆。在这种PC框架下,感官输入的记忆由Hebbian学习动力学[21]驱动,该动力学最小化输入和网络生成的预测之间的误差,并且通过推理神经动力学执行该输入的检索,也最小化内部预测之间的误差以及受损的感官输入。
这两种建模方法,即通过协方差学习的AM和通过预测处理的AM,看起来是完全不同和的,协方差学习模型忽略了海马活动的预测性质,而迄今为止用于模拟记忆任务的PCNs的架构仅包括神经元层(代表不同的皮质区域)之间的连接,因此从根本上不同于海马网络的已知连接,海马网络包括被认为在联想记忆中起关键作用的子场CA3中的循环连接[4]。这种二分法给从计算和理论的角度理解海马体带来了很大的困难。在这项工作中,我们旨在通过使用一族协方差学习PCN(cov PCN)统一建模AM的两种方法来解决这种二分法,并且在评估这些cov PCN时,我们特别关注数值稳定性和生物学合理性。具体来说,我们考虑以下标准[22]来验证我们模型的生物合理性:
1.局部计算:一个神经元的计算仅仅依赖于它的输入神经元和连接它自己和它的输入神经元的权重。
2.局部可塑性:一个模型中突触的可塑性规则只取决于其突触前和突触后神经元的乘积,即Hebbian学习规则[21]。
3.架构相似性:模型的组件类似于真实神经元的架构,例如海马体中锥体神经元的循环连接和顶端树突。
考虑到这些标准,我们首先发现早期的PCN已经整合了编码输入协方差信息的循环突触连接[23–25],因此可以认为是PC和协方差学习的统一。我们称之为显式covPCN,因为它将协方差矩阵显式编码到其递归突触中。显式covPCN最初是作为学习感觉输入表征的模型提出的,我们在这项工作中表明,它的协方差学习性质可以用于执行AM,因此它满足我们对海马中AM的单一模型的要求。然而,我们注意到,这个模型中协方差参数的学习规则是non-Hebbian的,因此是不可信的,并且也提出了重要的计算问题。为了解决这些问题,我们在这项工作中提出了一种新的递归PCN,它也通过其递归连接对协方差矩阵进行编码,但以隐式方式进行,因此我们将其称为隐式covPCN。我们证明了新的隐式模型也通过协方差学习执行AM,并且在简单的AM任务中,它在理论上和经验上都等价于显式covPCN,而只使用局部Hebbian可塑性。我们还表明,隐式模型可以进一步修改,通过引入树突结构实现与海马锥体细胞的生物学相似性,同时在收敛时保留与显式covPCN的理论和经验等效性。在这项工作中,我们将其命名为树枝状covPCN。重要的是,我们表明隐式和树状模型都可以执行更复杂的AM任务,其中显式covPCN会由于其不稳定的动力学而失败。最后,我们将隐式covPCN与分层PCN [19,20]结合起来,对整个海马-新皮质区进行建模,并表明它可以有效地执行具有挑战性的AM任务。
总的来说,我们的贡献是双重的:首先,我们在一个循环网络中实现了PC,这揭示了预测性处理是如何通过循环海马网络进行的。其次,我们证明了循环PCNs也保留了经典模型的协方差学习特性,从而将这些经典模型与PC框架统一起来。这种统一的描述极大地促进了对AM中海马执行的计算的理论理解
展望未来,由于 PC 与深度学习之间的密切关系,这种基于 PC 的模型有可能扩展到现代机器学习模型,这一点已被最近的研究 [22,26–28] 所揭示。因此,它的计算能力可能有助于对其他海马体功能进行建模,例如表征学习、导航和时间预测,从而指导未来对海马体执行的这些更复杂任务的计算和实验研究。



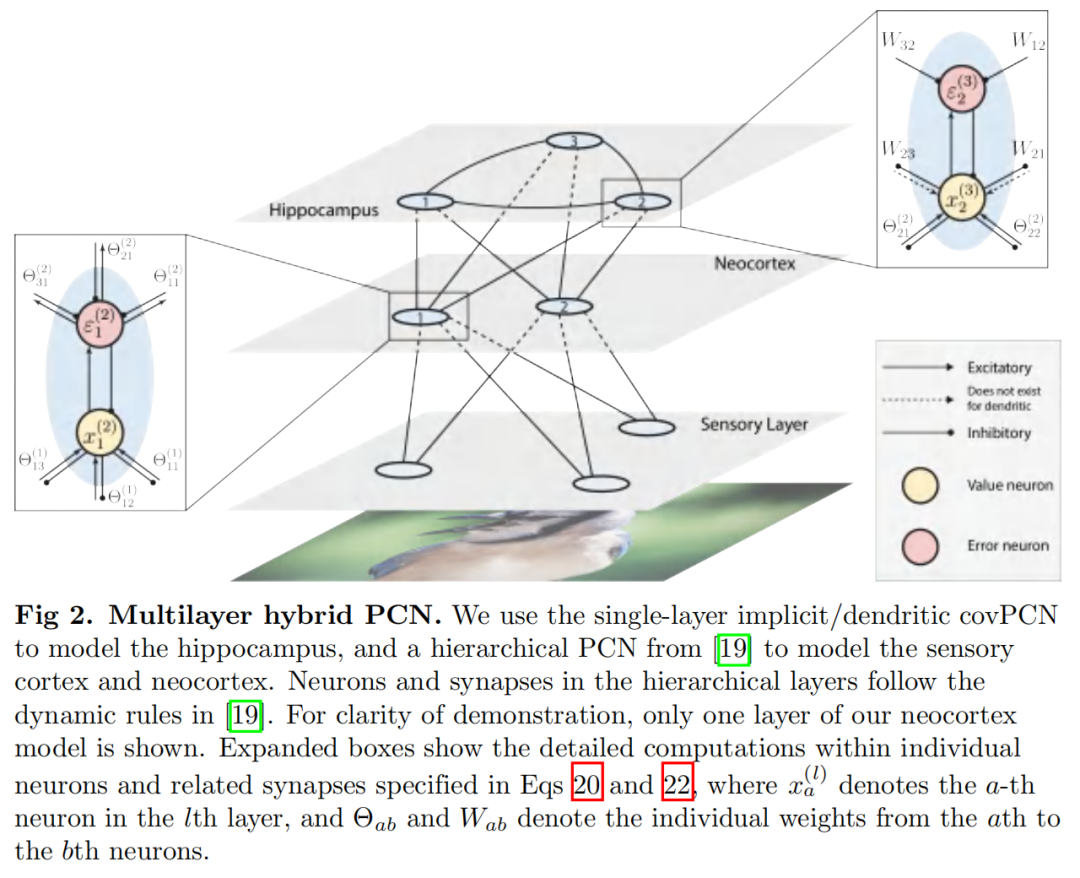
混合 PCN
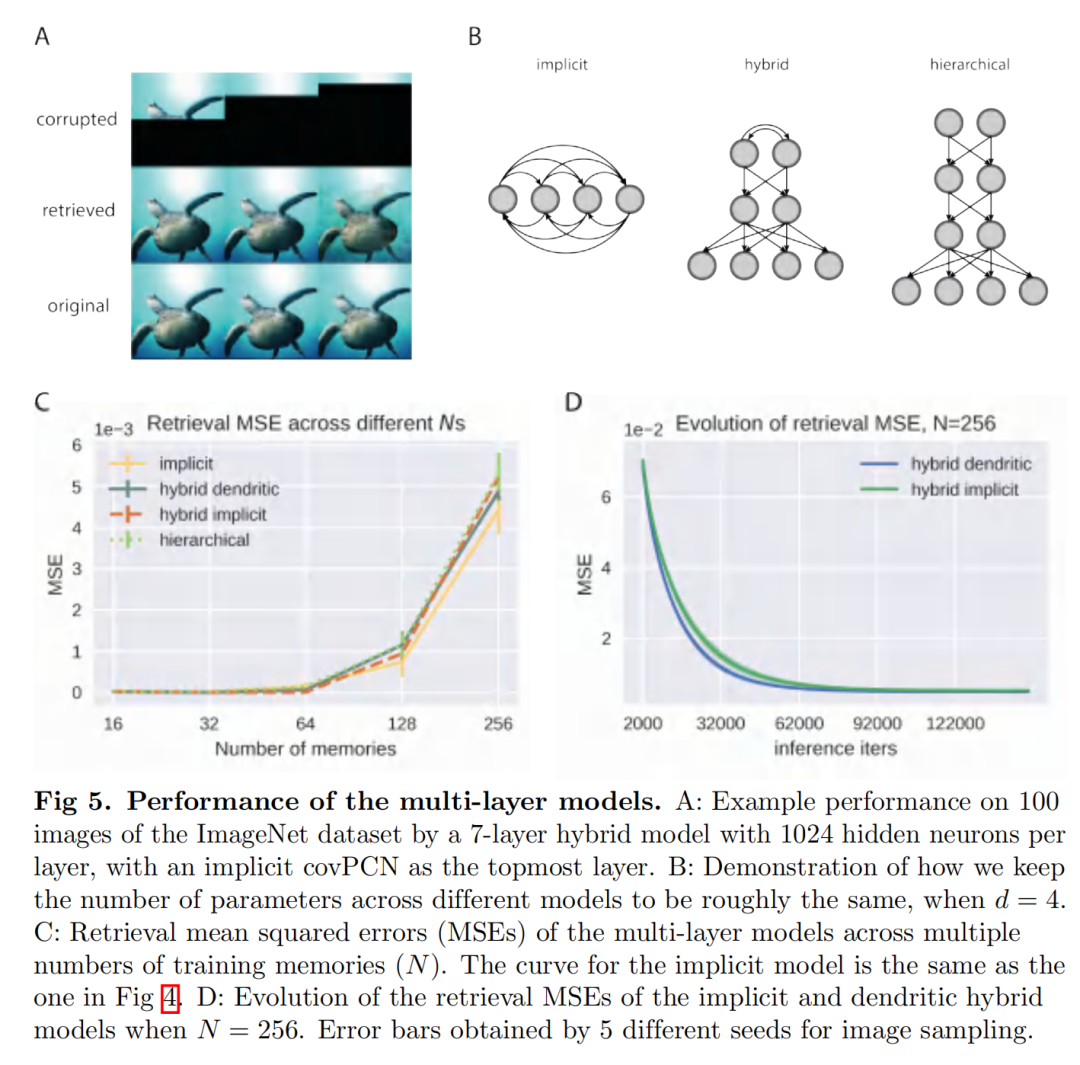
我们现在使用获得的结果提出一种架构,该架构模仿海马体的行为作为记忆索引和生成模型。上面介绍的循环单层网络模型为海马体中的循环动力学提供了模型。然而,原始感觉输入首先通过感觉层和新皮质层的层次结构进行处理,从而使海马体能够记住原始信号的表示,而不是信号本身。此外,根据最近的理论,海马体作为一种生成模型发挥作用,它累积来自层次结构较低的感觉和新皮质神经元的预测误差,并将下行预测发送到新皮质,以纠正新皮质神经元中的预测误差。海马体的生成模型不断更新,直到海马体预测通过抑制新皮层活动来纠正预测错误 [17]。分层 PCN 已经被提议作为一种生成模型来学习感官输入的表征 [24]。特别是,已经表明,没有任何循环结构的纯层次 PCN 能够在高度复杂的数据集上执行关联记忆任务 [19]。在这里,我们将分层 PCN 与我们提出的递归架构相结合,在最顶层获得具有递归动态的分层模型。结果是一个混合网络模拟了从感觉神经元到海马神经元的整个通路。
阅读原文参考完整原论文。
本文参与 腾讯云自媒体同步曝光计划,分享自微信公众号。
原始发表:2023/03/24 17:18:00,如有侵权请联系 cloudcommunity@tencent.com 删除
评论
登录后参与评论
推荐阅读
腾讯云开发者

Copyright © 2013 - 2026 Tencent Cloud. All Rights Reserved. 腾讯云 版权所有
深圳市腾讯计算机系统有限公司 ICP备案/许可证号:粤B2-20090059 ![]() 粤公网安备44030502008569号
粤公网安备44030502008569号
腾讯云计算(北京)有限责任公司 京ICP证150476号 | 京ICP备11018762号